Question
In: Chemistry
What kind of kinetics is observed in an enzymatic reaction, under conditions where the substrate concentration...
What kind of kinetics is observed in an enzymatic reaction, under conditions where the substrate concentration is low compared to the Km?
Solutions
Expert Solution
As enzyme-catalysed reactions are saturable, their rate of catalysis does not show a linear response to increasing substrate. If the initial rate of the reaction is measured over a range of substrate concentrations (denoted as [S]), the reaction rate (v) increases as [S] increases, as shown on the right. However, as [S] gets higher, the enzyme becomes saturated with substrate and the rate reaches Vmax, the enzyme's maximum rate.
The Michaelis–Menten kinetic model of a single-substrate
reaction is shown on the right. There is an initial bimolecular
reaction between the enzyme E and substrate S to form the
enzyme–substrate complex ES but the rate of enzymatic reaction
increases with the increase of the substrate concentration up to a
certain level but then an increase in substrate concentration does
not cause any increase in reaction rate as there no more E
available for reacting with S and the rate of reaction becomes
dependent on ES and the reaction becomes unimolecular reaction.
Although the enzymatic mechanism for the unimolecular reaction
 can be quite complex, there is typically one
rate-determining enzymatic step that allows this reaction to be
modelled as a single catalytic step with an apparent unimolecular
rate constant kcat. If the reaction path
proceeds over one or several intermediates,
kcat will be a function of several elementary
rate constants, whereas in the simplest case of a single elementary
reaction (e.g. no intermediates) it will be identical to the
elementary unimolecular rate constant k2. The
apparent unimolecular rate constant kcat is
also called turnover number and denotes the maximum number
of enzymatic reactions catalysed per second.
can be quite complex, there is typically one
rate-determining enzymatic step that allows this reaction to be
modelled as a single catalytic step with an apparent unimolecular
rate constant kcat. If the reaction path
proceeds over one or several intermediates,
kcat will be a function of several elementary
rate constants, whereas in the simplest case of a single elementary
reaction (e.g. no intermediates) it will be identical to the
elementary unimolecular rate constant k2. The
apparent unimolecular rate constant kcat is
also called turnover number and denotes the maximum number
of enzymatic reactions catalysed per second.
The Michaelis–Menten equation[9] describes how the (initial) reaction rate v0 depends on the position of the substrate-binding equilibrium and the rate constant k2.
 (Michaelis–Menten
equation)
(Michaelis–Menten
equation)
with the constants

This Michaelis–Menten equation is the basis for most
single-substrate enzyme kinetics. Two crucial assumptions underlie
this equation (apart from the general assumption about the
mechanism only involving no intermediate or product inhibition, and
there is no allostericity or cooperativity). The first assumption
is the so-called quasi-steady-state assumption (or
pseudo-steady-state hypothesis), namely that the concentration of
the substrate-bound enzyme (and hence also the unbound enzyme)
changes much more slowly than those of the product and substrate
and thus the change over time of the complex can be set to zero
 . The second assumption is that the total
enzyme concentration does not change over time, thus
. The second assumption is that the total
enzyme concentration does not change over time, thus  . A complete derivation can be found
here.
. A complete derivation can be found
here.
The Michaelis constant KM is experimentally
defined as the concentration at which the rate of the enzyme
reaction is half Vmax, which can be verified by
substituting [S] = Km into the Michaelis–Menten
equation and can also be seen graphically. If the rate-determining
enzymatic step is slow compared to substrate dissociation
(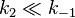 ), the Michaelis constant
KM is roughly the dissociation constant
KD of the ES complex.
), the Michaelis constant
KM is roughly the dissociation constant
KD of the ES complex.
If  is small compared to
is small compared to  then the term
then the term  and also very little ES complex is formed,
thus
and also very little ES complex is formed,
thus  . Therefore, the rate of product formation
is
. Therefore, the rate of product formation
is

Thus the product formation rate depends on the enzyme
concentration as well as on the substrate concentration, the
equation resembles a bimolecular reaction with a corresponding
pseudo-second order rate constant  . This constant is a measure of catalytic
efficiency. The most efficient enzymes reach a in the range of 108–
1010 M−1 s−1. These enzymes are so
efficient they effectively catalyse a reaction each time they
encounter a substrate molecule and have thus reached an upper
theoretical limit for efficiency (diffusion limit); and are
sometimes referred to as kinetically perfect enzymes
. This constant is a measure of catalytic
efficiency. The most efficient enzymes reach a in the range of 108–
1010 M−1 s−1. These enzymes are so
efficient they effectively catalyse a reaction each time they
encounter a substrate molecule and have thus reached an upper
theoretical limit for efficiency (diffusion limit); and are
sometimes referred to as kinetically perfect enzymes
 queen_honey_blossom answered 3 years ago
queen_honey_blossom answered 3 years agoRelated Solutions
An enzymatic reaction was analyzed for rate versus substrate concentration. Roughly estimate the Vmax and Km...
What assumption concerning substrate concentration is made in the discussion of enzyme kinetics? Why is this...
Wittig Reaction: Under what conditions does the Wittig reaction favor the cis isomer? Under what conditions does...
A student is given the task of determining the rate of an enzymatic reaction under defined...
1a) what is the relationship between substrate concentration and reaction rate? b) Describe how the enzyme...
4. What does enzymatic kinetics imply and how can we classify it? (Bibliography)
What are some enzymatic challenges used by plants to evolve under increasing co2 concentrations and where...
If the substrate concentration is limiting, how does increasing enzyme concentration affect reaction rate? A.Increasing enzyme...
What is anhedonia? In what conditions anhedonia is observed?
What is the Transition Reaction? What is the substrate? What are the products? What is Coenzyme A?
- The output of the function is a dictionary whose keys represent the bins and whose values...
- The shape of a graph of a binomial distribution depends on the value of both n...
- In 2012, cost per Medicare beneficiary did what?
- 3. A. What techniques can a firm use to optimize demand deposit holdings? B. How do...
- The half-life of mercury-197 is 64.1 hours. If a patient undergoing a kidney scan is given...
- Double bonds react with Br2 to form a dibromide. Isobutylene undergoes cationic polymerization under conditions where...
- 1. Which sex chromosomes are limited to only one sex? A. X and Z B. X...